墿
棏
妱
妱
尨嶕摦暔
歁擕椶
墿
棏

摍
妱
擃懱摦暔
椉惗椶

暘
妱
妱

唳拵椶
捁椶
墿
棏
妱
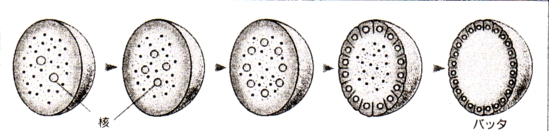
峛妅椶乯
仭惗暔
仭侽係擭搙俁妛婜妛擭枛峫嵏丂惗暔
丂堚揱巕昞尰朄偺婎杮偼傾儖僼傽儀僢僩偱偁傞丅 [ 桪惈 ] 堚揱巕偼戝暥帤丄
[ 楎惈 ] 堚揱巕偼彫暥帤偱昞婰偟丄彂偔嵺偵偼傾儖僼傽儀僢僩弴丄摨堦傾儖僼傽儀僢僩偺側偐偱偼戝暥帤偑愭偵側傞丅
丂AA傗aa側偳偺丄摨堦傾儖僼傽儀僢僩偱昞婰偝傟傞傕偺偼 [ 弮宯 ] 偲屇偽傟傞丅偙傟偼帺壠庴惛乮庴暡乯偟偰傕[
拲栚宍幙 ] 偑曄傢傞偙偲偼側偄丅堎側傞宍幙傪帩偮弮宯摨巑偺岎攝偵傛偭偰惗偠偨屄懱偼
[ 嶨庬 ] 偲屇偽傟傞丅屄懱偵偁傜傢傟傞宍幙偼 [ 昞尰 ] 宆丄屄懱偺帩偮堚揱巕偺峔惉偼
[ 堚揱巕 ] 宆偲屇偽傟傞丅傑偨丄昞尰宆偵偍偄偰憡懳偡傞宍幙傪 [ 懳棫宍幙
] 偲偄偆丅
丂愼怓懱偼 [ 嵶朎妀 ] 撪偵懚嵼偟丄 [ 堚揱巕乮DNA乯 ] 偲 [ 僞儞僷僋幙 ]
偐傜側傞丅懱嵶朎拞偵偼摨宍摨戝偺愼怓懱偑侾懳偢偮懚嵼偟偰偄傞丅偙偺侾懳偺堦曽偼晝曽丄堦曽偼曣曽偵桼棃偟偰偍傝丄
[ 憡摨愼怓懱 ] 偲屇偽傟傞丅攝嬼巕拞偺愼怓懱偼丄 [ 尭悢暘楐 ] 偵傛傝敿尭偟偰偄傞丅
丂侾慻偺懳棫宍幙偩偗偵拲栚偟偰嶌偭偨嶨庬偺偙偲傪 [ 堦堚揱巕嶨庬 ] 偲偄偆丅
丂弮宯偺懳棫宍幙傪岎嶨偡傞偲丄桪惈宍幙偺傒偑尰傟傞偙偲傪 [ 桪惈偺朄懃 ]
偲偄偄丄偙偺帪尰傟傞宍幙傪[ 桪惈宍幙 ] 丄尰傟側偄宍幙傪 [ 楎惈宍幙 ] 偲偄偆丅傑偨丄攝嬼巕宍惉帪偵堦懳偺堚揱巕偑暘偐傟丄暿乆偺攝嬼巕偵偼偄傞偙偲傪
[ 暘棧偺朄懃 ] 偲偄偆丅
丂AA丄aa側偳偺摨宆愙懕懱偺偙偲傪 [ 儂儌 ] 愙崌懱丄Aa側偳偺堎宆愙崌懱偺偙偲傪
[ 僿僥儘 ] 愙崌懱偲偄偆丅堚揱巕偑傢偐傜側偄桪惈宍幙屄懱偵 [ 楎惈 ] 堚揱巕偺
[ 儂儌 ] 愙崌懱傪岎嶨偟丄堚揱巕宆偲惗偢傞攝嬼巕斾傪挷傋傞偙偲傪 [ 専掕岎嶨
] 偲偄偆丅傑偨丄惗偠偨F1偲P傪岎嶨偡傞偙偲傪 [ 傕偳偟岎嶨 ] 偲偄偆丅
丂俀懳偺懳棫堚揱巕偵拝栚偟偰嶌偭偨嶨庬偺偙偲傪 [ 堚揱巕嶨庬 ] 偲偄偄丄昞尰宆丄堚揱巕宆傕俀庬椶傪慻傒崌傢偣偨傕偺偲側傞丅傑偨丄俀懳偺懳棫堚揱巕偑屳偄偵撈棫偟偰揱偊傜傟傞偙偲傪
[ 撈棫偺朄懃 ] 偲偄偆丅
丂懳棫堚揱巕偺桪楎娭學偑庛偄傕偺傪 [ 晄姰慡桪惈 ] 偲偄偆丅儅儖僶僫傾僒僈僆偺応崌丄R乮愒乯偺堚揱巕偲r乮敀乯偺堚揱巕偺桪楎娭學偑庛偄偨傔偵丄RR偲rr傪岎嶨偟偨Rr偼搷怓偲側傞丅傑偨丄Rr偼偙偺応崌桪楎拞娫偺宍幙偱偁傝丄
[ 拞娫嶨庬 ] 偲屇偽傟傞丅傑偨丄偙偺応崌堚揱巕宆偲昞尰宆偼堦抳偡傞丅
丂侾宍幙偵俁偮埲忋偺懳棫堚揱巕偑娭學偡傞傛偆側堚揱傪 [ 暋懳棫堚揱 ] 偲偄偆丅椺偲偟偰偼僸僩偺寣塼宆偺堚揱偑嫇偘傜傟傞丅偙偺応崌丄堚揱巕偼A丄B丄O偵暘偐傟丄
[ O ] 偵懳偟偰 [ A ] 丄 [ B ] 偼桪惈偩偑丄 [ A ] 偲 [ B ] 偵偼桪惈娭學偼側偄丅
丂儂儌愙崌懱偵側傞偙偲偵傛偭偰 [ 愭揤揑戙幱堎忢 ] 偑婲偒偨傝丄 [ 敪惗 ]
偑慾奞偝傟偨傝偡傞堚揱巕傪[ 抳巰堚揱巕 ] 偲偄偆丅偙傟偼懡悢偺惗暔偱抦傜傟偰偄傞丅椺偊偽丄栄怓偑墿怓偺堚揱巕Y偲敀怓偺堚揱巕y偱偼丄Y偑楎惈偺抳巰嶌梡傪帩偮偨傔偵惗傑傟傞墿怓屄懱偼偡傋偰Yy偲側傞丅
丂俀偮偺堚揱巕偑屳偄偵偦偺摦偒傪曔懆偟偁偭偰宍幙傪昞偡傕偺傪 [ 曗懌堚揱巕
] 偲偄偆丅椺偊偽僗僀乕僩僺乕偵偼怓慺尮傪嶌傞堚揱巕丄嶌傜側偄堚揱巕偺C丄c偲丄怓慺尮偐傜怓慺傪嶌傞堚揱巕丄嶌傜側偄堚揱巕偺P丄p偑偁傝丄C偲P偑摨帪偵懚嵼偟偰弶傔偰壴偑桳怓偲側傞丅
丂偁傞宍幙偺摥偒傪梷偊傞堚揱巕傪 [ 梷惂堚揱巕 ] 偲偄偆丅
丂帗梇堎懱偺惗暔偺応崌丄惈斾偼 [ 侾 ] 丗 [ 侾 ] 偱偁傞丅偙傟偼丄惈偵娭偡傞堚揱巕偺堦曽偑儂儌丄堦曽偑僿僥儘偺偨傔偱偁傞丅傑偨丄帗梇摨懱偺惗暔傗惈揮姺偡傞傕偺偼侾丗侾偱柍偄傕偺偑懡偄丅
丂惈寛掕偵娭梌偡傞愼怓懱偼 [ 惈愼怓懱 ] 偲屇偽傟傞丅傑偨丄帗梇偵嫟捠側愼怓懱偼
[ 忢愼怓懱 ] 偲屇偽傟傞丅侾慻偺忢愼怓懱傪A偲昞偡偲偡傞偲丄帗梇傪寛掕偡傞偺偼憡摨愼怓懱偺忢愼怓懱偺堦曽偺傒偱偁傞偐傜丄僸僩偺応崌俀n亖係俇偱丄亯偼
[ 俀俙亄倃倄 ] 丄亰偼 [ 俀俙亄倃倃 ] 偲側傞丅
丂X愼怓懱偵偁傞堚揱巕偼丄亯偺応崌楎惈偱傕昞傟傞偙偲偵側傞丅
丂堚揱巕偑偦傟偧傟撈棫偟偰偄傞応崌丄攝嬼巕偵偼偦傟偧傟偺堚揱巕偑擖傝 [
撈棫偺朄懃 ] 偑惉傝棫偮偑丄侾愼怓懱偵俀偮埲忋偺堚揱巕偑嫟桳偟偰偄傞応崌偼椉曽偲傕偑堦弿偵堚揱傪偡傞丅偙傟傪[
堚揱巕偺楢嵔 ] 偲偄偆丅
丂偁傞堚揱巕偑楢嵔偟偰偄偰傕丄偦偺楢嵔埲奜偺慻傒崌傢偣偑弌尰偡傞偲偒偑偁傞丅偙傟偼丄
[ 尭悢暘楐 ] 帪偵愼怓懱偺晹暘揑岎姺丄 [ 忔姺偊 ] 偲偄傢傟傞尰徾偑婲偒傞偐傜偱偁傞丅偙傟偵傛偭偰堦晹偺堚揱巕偑慻傒姺偊傜傟傞偙偲傪丄
[ 慻姺偊 ] 偲偄偆丅
丂F1乮僿僥儘乯傪専掕岎嶨偟偨偲偒偵惗偠傞F2偺昞尰宆暘棧斾偼丄 [ F1偺攝嬼巕 ] 偺暘棧斾偲摨偠偵側傞丅傑偨丄F2偺昞尰宆偺斾偑侾丗侾丗侾丗侾偵側傞偲偒偼 [ 撈棫偺朄懃 ] 偑惉傝棫偭偰偍傝丄側傜側偄偲偒偼
[ 楢嵔 ] 傪偟偰偄傞丅
丂堚揱巕慻傒姺偊偺婲偙傞昿搙偺偙偲傪慻姺偊斾偲偄偆丅偙傟偼丄
乮 [ 慻姺偊偺婲偙偭偨攝嬼巕偺悢 ] 乛 [ 攝嬼巕偺憤悢 ] 乯亊侾侽侽
偱媮傔傞偙偲偑弌棃傞丅
丂楢嵔偟偰偄傞堚揱巕偺僌儖乕僾偺偙偲傪 [ 楢嵔孮 ] 偲偄偆丅傑偨丄 [ 楢嵔孮
] 悢偼攝嬼巕拞偺 [ 愼怓懱悢 ] 偲堦抳偡傞丅
丂儌乕僈儞偼丄僉僀儘僔儑僂僕儑僂僶僄傪梡偄偰丄摨堦愼怓懱忋偺俁偮偺堚揱巕偺
[ 慻姺偊壙 ] 偐傜憡懳揑埵抲傪柧傜偐偵偡傞曽朄丄 [ 俁揰岎嶨 ] 傪偍偙側偭偨丅偦偟偰偙傟偵傛偭偰傢偐偭偨堚揱巕偺憡懳揑埵抲傪捈慄忋偵昞偟丄[
愼怓懱抧恾 ] 傪嶌惉偟偨丅偙偺抧恾偱偼丄楢嵔偡傞堚揱巕摨巑偺堚揱巕嫍棧偑戝偒偄傎偳慻姺偊偺婲偙傞棪偑[
崅 ] 偔側傝丄嫍棧偑彫偝偄傎偳慻姺偊偺婲偙傞棪偼 [ 掅 ] 偔側傞丅
丂憃憷椶偺偩態嵶朎偵忢帪傒傜傟傞 [ 偩態愼怓懱 ] 偼丄晛捠偺愼怓懱偺侾侽侽乣俀侽侽攞偺僒僀僘偱丄慡挿偵傢偨傝懡悢偺墶幦偑偁傞丅傑偨丄懠偵斾傋堎忢偵朿傜傫偩晹暘偼
[ 僷僼 ] 偲偄偄丄偙偙偱偼 [ RNA ] 偺揮幨偑峴傢傟偰偄傞丅
丒DNA偺敪尒
丂堚揱巕偺杮懱偼丄妀巁偺堦庬偱偁傞 [ DNA乮僨僆僉僔儕儃妀巁乯 ] 偱偁傞丅
丂DNA偼侾俉俇俋擭丄 [ 儊僢僸儍乕 ] 偵傛偭偰敪尒偝傟偨丅斵偼嵶朎偐傜DNA傪暘棧偟
[ 僰僋儗僀儞 ] 偲柤晅偗丄儕儞巁偵偲傫偩惈幙偵偮偄偰婰弎偟偨偑丄惗棟揑栶妱傗婡擻偵偮偄偰偼摉帪偼晄柧偺傑傑偩偭偨丅
丒堚揱巕偲僞儞僷僋幙乮峺慺乯偺娭學傪扵傞帋傒
丂侾俋侽俉擭丄 [ 僈儘乕僪 ] 偼 [ 僼僃儕僯働僩儞擜徢 ] 偺傛偆側堚揱惈戙幱堎忢傪
[ 峺慺寚懝徢 ] 偲娭楢偯偗丄偝傜偵堚揱妛揑側崻嫆傪帩偮偲怣偠偨丅
丂侾俋侽俋擭偵偼 [ 僀僃儞僙儞 ] 偑堚揱揑宍幙偺揱払偵娭梌偡傞暔幙傪堚揱巕偲柤晅偗偨丅
丂侾俋係侾擭丄 [ 價乕僪儖仌僥乕僞儉 ] 偼傾僇僷儞僇價偵X慄傪徠幩丄撍慠曄堎傪婲偙偝偣偰條乆側
[ 塰梴揑寚娮 ] 偺偁傞宯摑丄偡側傢偪惗堢偡傞偨傔偵摿掕偺 [ 戙幱暔幙 ] 偑昁梫偲側傞庬傪嶌偭偨丅偦偟偰丄撍慠堎曄庬摨巑傪岎嶨偟丄峺慺寚懝徢偑
[ 儊儞僨儖堚揱 ] 妛偵婎偯偔偙偲傪妋擣丄堚揱巕偲峺慺偺娫偺娭學傪妋棫偟偨丅[
價乕僪儖 ] 偼傑偨丄侾俋俆侾擭偵 [ 堦堚揱巕堦峺慺 ] 愢傪採彞偟偰偄傞丅
丒懱嵶朎偲攝嬼巕偵偍偗傞DNA偺検揑娭學
丂侾俋係俉丄係俋擭偵丄儈儖僗僉乕傜偼愼怓懱偺俀攞懱乮俀値乯偁偨傝偺DNA検偼惗暔偺庬偵傛傝堦掕偱偁傝丄偦傟偼[
惛巕嵶朎 ] 偵偁傞DNA検偺俀攞偱偁傞偙偲傪帵偟偨丅傑偨偙傟偼丄懠幰偺抦尒傛傝乽DNA偼戙幱揑偵埨掕暔幙偱丄愼怓懱偵懡偔娷傑傟丄娙扨偵
[ 崌惉丒暘夝 ] 偝傟偢丄嵶朎偑 [ 寬慡 ] 側傜偽偦偙偐傜棧傟側偄乿偲偄偆忣曬偲崌傢偣傞偲丄DNA偑堚揱暔幙偱偁傞偙偲偺埫帵偲側傞丅
丒DNA偺婡擻丄堚揱巕偺杮懱偼DNA偱偁傞偙偲偺徹嫆
丂侾俋俀俉擭丄 [ 僌儕僼傿僗 ] 偼撆惈偺側偄宯摑偺嵶朎桼棃偺惗嵶朎偑丄桳撆偱攛墛傪堷偒婲偙偡宯摑偺嵶朎偲崿崌偝傟傞偲
[ 宍幙揮姺 ] 傪婲偙偡偙偲傪敪尒偟偨丅偟偐偟丄壗偵傛偭偰宍幙揮姺偑婲偙傞偺偐傑偱偼暘偐傜側偐偭偨丅
丂偦偺屻偺侾俋係係擭丄 [ 僄僀僽儕乕 ] 傜偼撆惈偺偁傞嵶朎桼棃偺巰傫偩嵟嬤嵶朎偺戙傢傝偵丄嶦偟偨嵶嬠偺拪弌暔幙偲偟偰惛惢偝傟偨DNA傪巊偄丄僌儕僼傿僗偲摨條偺寢壥傪摼偨丅偙傟偵傛傝宍幙揮姺偼DNA偵傛偭偰婲偙偭偨傕偺偱偁傝DNA偑堚揱擻椡傪帩偮偲寢榑丅堚揱偲DNA偺娭學偑柧妋偲側偭偨丅丂侾俋俆俀擭丄
[ 僴乕僔乕仌僠僃僀僗 ] 偼 [ 僶僋僥儕僆僼傽乕僕 ] 偑廻庡偺嵟嬤嵶朎偵堚揱暔幙傪拲擖偡傞偙偲傪抦偭偨丅偙傟偵傛傝丄僂僀儖僗偺堚揱巕傕DNA偱偁傞偙偲偑撍偒偲傔傜傟偨丅
丂偟偐偟丄偙偺帪揰偱偼傑偩DNA偑偳偺傛偆側棫懱峔憿偱丄堚揱巕暔幙偺敪尰傪偡傞偺偐摍偼晄柧偱偁偭偨丅
丂妀巁偼丄傎偲傫偳偑 [ 妀 ] 偵懚嵼偟 [ 擇杮嵔 ] 峔憿偱偁傞DNA乮僨僆僉僔儕儃妀巁乯偲丄
[ 妀 ] 丒[ 嵶朎幙婎幙 ] 偵懚嵼偟 [ 堦杮嵔 ] 峔憿偱偁傞RNA乮儕儃妀巁乯偲偵暘椶偱偒傞丅傑偨丄妀巁偺婎杮扨埵傪[
僰僋儗僆僠僪 ] 偲偄偆丅DNA偺応崌丄僰僋儗僆僠僪偼 [ 儕儞巁 ] 丄 [ 摐乮僨僆僉僔儕儃乕僗乯
] 丄 [ 墫婎 ] 偐傜峔惉偝傟偰偄傞丅傑偨丄墫婎偼A乮 [ 傾僨僯儞 ] 乯丄T乮
[ 僠儈儞 ] 乯丄G乮 [ 僌傾僯儞 ] 乯丄C乮 [ 僔僩僔儞 ] 乯丄偺巐庬椶偑偁傞丅
丂侾俋係俋擭丄 [ 僔儍儖僈僼 ] 偼DNA拞偺係墫婎偺検揑娭學傪挷傋丄 [ A ] 佮
[ T ] 丄 [ G ] 佮 [ C ] 偲偄偆検揑娭學偵側傞偙偲傪敪尒偟偨丅偙傟傪 [ 僔儍儖僈僼偺朄懃
] 偲偄偆丅
丂傑偨丄X慄寢徎妛偺尋媶幰偱偁傞 [ 僂傿儖僉儞僗仌僼儔儞僋儕儞 ] 偼丄侾俋俆俀丆俆俁擭偵DNA寢徎偵X慄傪摉偰丄峔憿傪挷傋偨丅偦偺寢壥丄DNA偑
[ 俀廳傜偣傫 ] 峔憿偱偁傞偙偲傪柧傜偐偵偟偨丅傑偨丄偙偺帪孞傝曉偟弌偰偔傞暘巕娫嫍棧偺抣丄
[ 0,34nm ] 丄 [ 2.0nm ] 丄 [ 3.4nm ] 偑偁偭偨偑丄偙傟偑昞偡偺偑壗側偺偐偼晄柧偱偁偭偨丅
丂偦偟偰侾俋俆俁擭丄 [ 儚僩僜儞仌僋儕僢僋 ] 偼DNA峔憿偵娭偡傞擖庤壜擻側忣曬偐傜丄DNA暘巕峔憿傪寛掕丄僲乕儀儖徿傪庴徿偟偨丅偙偺暘巕峔憿儌僨儖偼
[ 儚僩僜儞丒僋儕僢僋儌僨儖 ] 偲屇偽傟傞丅
偙傟偵傛傞偲丄 [ DNA暘巕暆 ] 偼2.0nm丄 [ 墫婎懳 ] 偼0.34nm偱丄 [ 傜偣傫侾夞揮
] 偺挿偝偑3.4nm偱偁偭偨丅傑偨丄 [ 僔儍儖僈僼偺朄懃 ] 傛傝A偼T丄G偼C偲寢崌偡傞偙偲傪柧傜偐偵偟丄俀廳傜偣傫偺峔憿傪
[ 摐 ] 丄[ 儕儞巁 ] 傛傝側傞俀杮偺傜偣傫忬嵔偑 [ 墫婎 ] 偱寢崌偟偨峔憿偱偁傞偲偟偨丅堦曽偺嵔偺僰僋儗僆僠僪偼懠曽偵懳偟偰媡岦偒偱偁傝丄庬偛偲偺DNA偺堘偄偼
[ 墫婎攝楍 ] 偺堘偄偱偁傞偲掕媊偟偨丅偪側傒偵僸僩偺寢崌偟偨僰僋儗僆僠僪偼栺
[ 俁侽壄 ] 偱丄堚揱巕検偼 [ 俁俆侽侽侽 ] 掱搙偱偁傞丅
丂DNA偑 [ 僞儞僷僋幙乮僸僗僩儞乯 ] 偵傑偒偮偒丄悢抜奒偺 [ 僐僀儖忬 ] 峔憿傪傊偰嬅廤偟偨傕偺偑丄崱擔偱偼愼怓懱偲屇偽傟傞傕偺偺惓懱偱偁傞丅
仭侽係擭搙俀妛婜婜枛峫嵏
丂庴梕婍偲偼丄奜奅偐傜偺巋寖傪庴偗庢傝丄嫽暠偲偟偰擼偵揱偊傞傕偺偱偁傞丅偙傟偼晀姶偵嫽暠偡傞
[ 姶妎嵶朎 ] 偑廤崌偟偰偱偒偰偄傞丅
丂恄宱偼丄 [ 拞悤 ] 恄宱宯偲 [ 枛徑 ] 恄宱宯偵暘偐傟傞丅拞悤恄宱宯偼 [
擼 ] 丄 [ 愐悜 ] 側偳偱丄枛徑恄宱宯偼偝傜偵 [ 姶妎 ] 恄宱丄 [ 塣摦 ] 恄宱側偳偺
[ 懱惈 ] 恄宱偲丄 [ 岎姶 ] 恄宱丄 [ 暃岎姶 ] 恄宱側偳偺 [ 帺棩 ] 恄宱宯偵暘偗傞偙偲偑偱偒傞丅
丂岠壥婍偲偼 [ 嬝擏 ] 傗 [ 態 ] 偺偙偲偱丄 [ 拞悤 ] 恄宱偐傜偺嫽暠偱斀墳偡傞丅堦斒偵丄嵶朎偑巋寖傪庴偗傞偲嵶朎枌偵
[ 揹婥 ] 揑曄壔偑敪惗偟丄妶摦忬懺偵側傞丅偙傟傪嵶朎偑嫽暠偡傞偲偄偆丅庴梕婍偱姶妎嵶朎偑庴梕偱偒傞巋寖偺庬椶傪
[ 揔巋寖 ] 偲偄偆丅椺偊偽丄娽偼岝偑揔巋寖偱偁傝丄岝傪庴怣偡傞偙偲偵傛偭偰帇妎偲偟偰懆偊傞偙偲偑弌棃傞偑丄壒傗廳椡傪娭抦偡傞偙偲偼弌棃側偄丅傑偨丄
[ 揔巋寖 ] 偵傛偭偰姶妎嵶朎偑嫽暠偡傞偨傔偵昁梫側巋寖偺嵟彫抣傪 [ 鑷抣
] 偲偄偆丅
丂岝偺巋寖偵傛偭偰惗偢傞姶妎傪帇妎偲偄偆丅岝傪姶偠傞偺偼 [ 帇 ] 嵶朎偱偁傞丅偙傟偼摿掕偺
[ 怓慺 ] 傪帩偪丄岝偱暘夝偝傟傞偙偲偵傛偭偰嫽暠傪惗偠傞丅
丂帇嵶朎偵偼 [ 悕懱 ] 嵶朎偲 [ 瀰懱 ] 嵶朎偑偁傞丅
丂[ 悕懱 ] 嵶朎偼庡偵柧強偱摥偔嵶朎偱偁傝丄栐枌拞怱晅嬤偺 [ 墿斄 ] 偵懡偄丅愒丄惵丄椢怓偵偦傟偧傟斀墳偟嫽暠偡傞嵶朎偑偁傝丄暔懱偺
[ 怓 ] 傗 [ 宍 ] 傪偼偭偒傝偲姶妎偡傞偑丄庛岝偱偼摥偐側偄偨傔偵丄埫強偱
[ 怓 ] 幆暿偑
偱偒側偄丅
丂[ 瀰懱 ] 嵶朎偼栐枌廃曈偵懡偔丄 [ 庛岝 ] 偵懳偟偰嫽暠偡傞丅傑偨丄 [ 怓姶妎
] 偵偼娭梌偟偰偄側偄丅
丂[ 栍斄 ] 偵偼帇嵶朎偼暘晍偟偰偄側偄丅
丂埫強偐傜柧強偵堏傞偲偒丄峥偟偄偲姶偠偰偐傜彊乆偵惓忢偵側偭偰偄偔丅偙傟傪
[ 柧弴墳 ] 偲偄偄丄峥偟偄偲姶偠傞偺偼[ 瀰懱 ] 嵶朎偑夁搙偵嫽暠偟偰偄傞偨傔偱偁傞丅媡偵柧強偐傜埫強偵堏傞偲偒丄偼偠傔偼壗傕尒偊側偄偑偟偽傜偔偡傞偲尒偊傞傛偆偵側傞丅偙傟傪
[ 埫弴墳 ] 偲偄偆丅偟偽傜偔偡傞偲尒偊傞傛偆偵側傞偺偼丄 [ 瀰懱 ] 嵶朎偑彊乆偵婡擻偟偼偠傔丄師戞偵庛岝傪姶偠傞偨傔偱偁傞丅岝検挷愡偼
[ 擑嵤 ] 偱峴側傢傟傞丅
丂僸僩偺娽偺墦嬤挷愡偼 [ 悈徎懱 ] 偺岤偝傪曄偊傞偙偲偵傛偭偰挷愡偝傟傞丅嬤偔傪尒傞偲偒偼栄條懱偑
[ 廂弅 ] 偟丄[ 僠儞彫懱 ] 偑備傞傓偙偲偵傛偭偰悈徎懱偑 [ 岤偔 ] 側傝丄墦偔傪尒傞偲偒偼栄條懱偑
[ 抩娚 ] 偟偰僠儞彫懱偑[ 堷偭挘傞 ] 偙偲偵傛偭偰悈徎懱偑 [ 敄偔 ] 側傞丅
丂嬻婥偺怳摦偵傛傞壒攇巋寖偵傛偭偰婲偙傞巋寖傪挳妎偲偄偆丅嬻婥怳摦偵傛傞壒攇偼
[ 帹妅 ] 仺 [ 奜帹摴 ] 仺 [ 屰枌 ] 仺 [ 帹彫崪 ] 仺乮 [ 棏墌憢 ] 乯仺撪帹偺
[ 偆偢傑偒娗 ]偺儕儞僷塼乮 [ 慜掚奒 ] 仺 [ 屰幒奒 ] 乯仺偆偢傑偒娗偺 [
婎掙晹 ] 仺 [ 僐儖僠婍挳 ] 嵶朎仺 [ 挳恄宱 ] 仺戝擼乮挳妎乯偺弴偵揱傢傞丅撪帹儕儞僷塼偑怳摦偡傞偲丄
[ 婎掙枌 ] 傕怳摦偟丄 [ 婎掙枌 ] 忋偺 [ 僐儖僠婍偍偍偄 ] 枌偵怗傟偰偄傞[
姶妎栄 ] 偑巋寖偝傟丄挳嵶朎偑嫽暠偡傞丅
丂廳椡丄塣摦曽岦傪庴梕偟偰惗偠傞姶妎傪暯峵妎偲偄偆丅歁擕椶偺応崌丄撪帹偺
[ 慜掚 ] 偲 [ 敿婯娗 ] 偑偦偺栶妱傪壥偨偟偰偄傞丅廳椡曽岦偲偦偺曄壔偼 [
慜掚 ] 偺姶妎嵶朎偲 [ 帹愇 ] 偺僘儗偵傛偭偰姶偠丄夞揮塣摦曽岦傗懍偝偼[
敿婯娗婎晹 ] 偺姶妎嵶朎偺 [ 姶妎栄 ] 偑 [ 儕儞僷塼 ] 偺棳傟傪姶抦偟偰姶偠傞丅歬妎婍偼歬嵶朎丄枴妎婍偼枴嵶朎偐傜偦傟偧傟弌棃偰偄傞丅
丂惗暔恄宱偼 [ 僯儏乕儘儞 ] 偑峔憿偺婎杮扨埵偱偁傞丅 [ 僯儏乕儘儞 ] 偼
[ 妀 ] 偲偦偺廃曈偺嵶朎懱丄 [ 幉嶕 ] 丄[ 庽忬撍婲 ] 偐傜側傞丅僯儏乕儘儞偼戝偒偔暘偗傞偲俀偮偺庬椶偵暘偗傜傟傞丅
[ 柍悜恄宱 ] 慇堐偼 [ 悜忊 ] 偑柍偔丄[ 柍愐捙 ] 摦暔偺恄宱慡斒傗丄 [ 愐捙
] 摦暔偺 [ 岎姶 ] 恄宱偑偙傟偵偁偨傞丅 [ 桳悜恄宱 ] 慇堐偼 [ 悜忊 ] 偑偁傝丄
[ 愐捙 ] 摦暔偺懡偔偺恄宱偑偙傟偵偁偨傞丅
丂恄宱慇堐偺枛抂偺晹暘偼 [ 僔僫僾僗 ] 偲屇偽傟傞丅僯儏乕儘儞摨巑偼傢偢偐側嫍棧傪偍偄偰愙懕偟偰偍傝丄僯儏乕儘儞偐傜僯儏乕儘儞偵嫽暠傪揱払偡傞栶妱傪帩偭偰偄傞丅
丂惷巭忬懺偺恄宱嵶朎偼丄幉嶕撪偺 [ 僀僆儞岎姺億儞僾 ] 偑 [ 擻摦 ] 桝憲偵傛偭偰
[ Na+ ] 傪嵶朎奜傊偲攔弌偟偰偄傞偨傔丄嵶朎枌偺撪奜偱僀僆儞暘晍偵嵎偑偁傞忬懺偵側偭偰偄傞丅偙偺嵎偼嵶朎偺撪偲奜偱惗偢傞揹埵偺嵎偲側傝丄奜懁偺揹埵偑崅偔側傞乮奜亄丒撪亅乯丅偙偺條偵嵶朎偺撪奜偱堦掕偺揹埵嵎偑曐偨傟偰偄傞忬懺傪
[ 惷巭揹埵 ] 偲偄偆丅嵶朎枌偑巋寖傪姶偠傞偲丄偦偺晹埵偵Na+偑棳擖偟丄揹埵偑媡揮乮奜亅丒撪亄乯丄嵶朎偑嫽暠偡傞丅嵶朎偺嫽暠偵敽偭偰惗偠傞揹埵傪
[ 妶摦揹埵 ] 偲偄偆丅堦搙嫽暠偟偨嵶朎偼偟偽傜偔偡傞偲尦偺忬懺偵栠傝丄偟偽傜偔偺娫晄妶惈壔偟偰[
愨懳晄擻 ] 婜偵側傞丅嵶朎偺嫽暠偼嫽暠晹偲椬愙偟偨椉懁偵惗偠傞揹埵嵎偵傛偭偰椉椬傊偲揱払偝傟偰偄偔偑丄堦搙嫽暠偟偨嵶朎偼晄妶惈壔偟偰偄傞偨傔偵嫽暠偑媡棳偡傞偙偲偼側偄丅偦偺偨傔丄嫽暠晹埵偱偼嫽暠偼椉曽岦偵揱払偝傟丄偦傟埲奜偱偼堦曽岦偵恑傓偺偩丅
丂丂丂丂 丂丂亄亄亄亄亄亄 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂亄亄亅亄亄亄 丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂 亄亅亄亅亄亄
丂丂丂丂劅劅劅劅劅劅劅劅劅劅丂仼嵶朎枌丂丂丂丂劅劅劅劅劅劅劅劅劅劅丂丂丂丂丂丂丂丂丂丂丂劅劅劅劅劅劅劅劅劅劅
丂丂丂丂丂 丂亅亅亅亅亅亅 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂亅亅亄亅亅亅 丂丂丂丂丂丂丂丂丂 丂丂丂丂丂 丂亅亄亅亄亅亅
丂丂丂丂丂丂丂丂幉嶕撪晹丂丂丂丂丂丂丂丂丂仺丂丂丂丂丂丂丂丂丂丂 嫽暠丂丂丂丂丂丂丂丂丂丂丂仺丂丂丂丂丂丂 丂丂 嫽暠揱払
丂丂丂丂丂丂 亅亅亅亅亅亅 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂亅亅亄亅亅亅 丂丂丂丂丂丂丂丂丂 丂丂丂丂 丂丂亅亄亅亄亅亅
丂丂丂丂劅劅劅劅劅劅劅劅劅劅丂丂丂丂丂丂丂丂丂丂丂劅劅劅劅劅劅劅劅劅劅丂丂丂丂丂丂丂丂丂丂丂劅劅劅劅劅劅劅劅劅劅
丂丂丂丂丂 丂亄亄亄亄亄亄 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂亄亄亅亄亄亄 丂丂丂丂丂丂丂丂丂丂 丂丂丂 丂丂亄亅亄亅亄亄
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂
丂 晄妶惈忬懺
丂桳悜恄宱慇堐偼丄愨墢懱偱偁傞悜忊偵曪傑傟偰偄傞偑丄悜忊偺愗傟栚偱偁傞
[ 儔儞價僄峣椫 ] 傪挼桇偡傞傛偆偵揹棳偑棳傟傞偨傔偵揹棳偺揱払懍搙偑懍偔丄
[ 100m/昩 ] 傎偳偱揹棳傪揱払偡傞丅柍悜恄宱慇堐偼悜忊偑側偄偨傔丄[ 悢m/昩
] 偲丄揱払懍搙偼抶偄丅妶摦揹埵偼巋寖偺戝偒偝偑堦掕乮鑷抣乯埲忋偵払偟側偄偲惗偠側偄偑丄鑷抣埲忋偱偁傟偽巋寖偺戝偒偝偵娭學側偔妶摦揹埵偼堦掕偱偁傞丅偙傟傪
[ 慡偐柍偐 ] 偺朄懃偲偄偆丅
丂嫽暠偑恄宱慇堐枛抂偺僯儏乕儘儞傑偱摓払偡傞偲丄僯儏乕儘儞偼撪晹偵偁傞
[ 僔僫僾僗彫朎 ] 偐傜[ 僲儖傾僪儗僫儕儞 ] 傗[ 傾僙僠儖僐儕儞 ] 側偳偺恄宱揱払暔幙傪曻弌偟丄椬愙偡傞僯儏乕儘儞偺
[ 庽忬撍婲 ] 傗[ 嵶朎枌 ] 傪巋寖偟偰嫽暠傪揱払偡傞丅
丂恄宱偼 [ 尨惗惗暔 ] 丄 [ 奀柸惗暔 ] 埲奜偺摦暔慡偰偵懚嵼偟偰偄傞丅扨嵶朎惗暔偺孮懱僞僀僾偼屄懱亖嵶朎偑偦傟偧傟偵巋寖傪庴偗偰偄傞偺偵懳偟丄懡嵶朎惗暔偺応崌偼恄宱偵傛偭偰嵶朎傪僱僢僩儚乕僋壔偟丄忣曬偺嫟桳丄摑崌壔偑側偝傟偰偄傞丅
丂 [ 嶶嵼 ] 恄宱宯偲偼丄懱慡懱偵恄宱偑栐栚忬偵暘晍偟偰偄傞庬椶偺偙偲偱丄
[ 僸僪儔 ] 傗 [ 僋儔僎 ] 側偳偑偙傟偵偁偨傞丅
丂 [ 偐偛宆 ] 恄宱宯偼摢晹偵 [ 恄宱愡 ] 偲偄偆嵶朎懱偺廤崌偟偨傕偺偑偁傞庬椶偱丄
[ 僾儔僫儕傾 ] 側偳偺[ 傊傫宍 ] 摦暔偑偙傟偵偁偨傞丅
丂 [ 偼偟偛宆 ] 恄宱宯偼懱愡枅偵恄宱愡偑 [ 侾 ] 懳偁傞庬椶偱丄 [ 娐宍 ]
摦暔傗 [ 愡懌 ] 摦暔偵尒傜傟傞丅[ 擃懱 ] 摦暔偼懱愡偑側偄偨傔丄懱奺晹偵恄宱愡偑懚嵼偡傞丅
丂 [ 娗忬 ] 恄宱宯偼 [ 擼 ] 丄 [ 愐悜 ] 偑拞悤偱偁傞丅偙傟傜偼敪惗夁掱偱
[ 恄宱娗 ] 偐傜暘壔偟偨傕偺偱偁傞丅[ 愐捙 ] 摦暔偑偙傟偵偁偨傞丅
丂愐悜偺抐柺傪尒傞偲奜懁偲撪懁偲偱堎側傞峔惉傪偟偰偄傞偙偲偑暘偐傞丅奜懁偼
[ 敀幙 ] 偲屇偽傟丄 [ 恄宱慇堐 ] 偺懇偱弌棃偰偄傞丅撪懁偼 [ 奃敀幙 ] 偲屇偽傟丄嵶朎懱偑懡偄丅嫽暠偑戝擼偵揱傢傝姶妎傪惗偠傞慜偵嬝擏偵嫽暠偑揱傢傞偙偲傪
[ 斀幩 ] 偲偄偆丅偙傟偼柍堄幆揑偵峴傢傟丄斀幩偑婲偒傞嵺偵嫽暠偺揱傢傞宱楬偺偙偲傪
[ 斀幩媩 ] 偲偄偆丅 [ 孅嬝 ] 斀幩傗 [ 旼奧寬 ] 斀幩側偳偼愐悜斀幩偺堦庬偱偁傞丅
丂戝擼偼嵍塃敿媴偑偁傝丄嵍塃偱摥偒偑堘偆丅戝擼旂幙偼 [ 奃敀幙 ] 偱弌棃偰偍傝丄嵶朎懱偑懡偄丅昞柺偺怴旂幙偼[
姶妎 ] 拞悤丄[ 悘堄塣摦 ] 拞悤丄 [ 婰壇 ] 丄 [ 巚峫 ] 丄 [ 棟夝 ] 側偳傪峴偆
[ 惛恄妶摦 ] 拞悤傪巌偭偰偄傞丅怴旂幙埲奜偺媽旂幙偱偼 [ 杮擻峴摦 ] 傪巌偭偰偄傞丅
丂撪懁偼 [ 戝擼悜幙 ] 偲屇偽傟丄恄宱慇堐偑廤崌偟偨 [ 敀幙 ] 偐傜側傞丅
丂彫擼偼 [ 塣摦 ] 偺挷愡傗懱偺 [ 暯峵 ] 傪曐偮婡擻傪巌偭偰偍傝丄 [ 悈拞
] 丄 [ 嬻拞 ] 偱妶摦偡傞摦暔偺応崌偼丄摿偵敪払偑寖偟偄丅
丂娫擼偼嫽暠傪愐悜偐傜戝擼傊偲揱偊傞 [ 帇彴 ] 丄 [ 帺棩恄宱 ] 拞悤偱偁傞
[ 帇彴壓晹 ] 偐傜側傞丅懠偵傕[ 懱壏 ] 丄[ 悈暘 ] 丄 [ 寣埑 ] 偺挷愡傪峴偆拞悤偱傕偁傞丅拞擼偼
[ 娫擼 ] 偐傜 [ 彫擼 ] 傊偺楢棈捠楬偲側偭偰偄傞丅墑悜偼惗柦堐帩偵捈愙娭學偟丄
[ 屇媧 ] 丄 [ 怱憻柆攺 ] 丄 [ 徚壔娗妶摦 ] 丄[徚壔塼暘斿 ] 側偳傪巌傞丅娫擼丄拞擼丄墑悜偺俁偮偼崌傢偣偰
[ 擼姴 ] 偲屇偽傟傞丅
丂枛徑恄宱宯偼丄擼偐傜弌偰偄傞 [ 侾俀 ] 懳偺擼恄宱丄愐悜偐傜弌偰偄傞 [
俁侾 ] 懳偺愐悜恄宱偵暘偐傟傞丅
仭侽係擭搙俀妛婜拞娫峫嵏
摦暔偺敪惗
敪惗丗柍惈揑偵惗偠偨夎丄朎巕側偳傪弌敪揰偲偟偰偦傟偑惉懱偵摓払偡傞夁掱乧庴惛棏側偳
敪惗偺抜奒
丒惉挿乧嵶朎暘楐丗敪惗弶婜偺懱嵶朎暘楐仺棏妱丄偦傟偵傛偭偰惗偠傞嵶朎妱媴
丂丂乥丂 丒妱媴偼戝偒偔側傜側偄仺堦斒偺懱嵶朎暘楐傛傝傕懍偄暘楐
丂丂乥
丂丂伀
丒嵶朎暘楐乧憹壛偟偨嵶朎偑偩傫偩傫偲堎側偭偨嵶朎偵側偭偰偄偔夁掱
丂丂乥丂丂丂丂乮怴婡擻丄壔妛揑峔憿柺偱偩傫偩傫偲摿庩壔偝傟傞夁掱乯
丂丂乥丂丂丂 丒暘巕仺堚揱巕偺敪尰乧堎暘巕宍惉傊
丂丂乥丂丂丂劇劅仺堦斒揑側懡嵶朎摦暔嵶朎乧晄壜媡揑側曄壔乮尦偵偼栠傟側偄乯
丂丂乥丂丂丂劎
丂丂乥丂丂丂劋劅仺崅摍怉暔嵶朎乧壜媡揑側曄壔仺姰慡嵞惗乮嵞暘壔乯偑壜擻
丂丂乥
丂丂伀
丒宍懺宍惉乧峔憿偲偟偰擣幆偱偒傞宍偑弌尰偟敪払偡傞夁掱
丂丂丂丂丂丂丂丂乮僾儘僌儔儉偝傟偨嵶朎巰傕娷傑傟傞仺椺丗僆僞儅僕儍僋僔偺偟偭傐乯
棏偺庬椶偲棏妱
丒懡嵶朎摦暔棑偺柤徧
丂丂 丂劇劅仺摦暔嬌乧廳椡偺偐偐傞斀懳懁偱丄嬌懱偺惗偠傞懁
丂棏丂劎丂丂丂丂丂丂丂丂丂偙偺嬌偺敿媴懁乧摦暔敿媴
丂丂 丂劋劅仺怉暔嬌乧摦暔嬌偵懳棫偡傞嬌
丂丂丂丂丂丂丂丂丂丂丂丂丂丂偙偺嬌偺敿媴懁乧怉暔敿媴
丒棏偺庬椶偲棏妱
| 棏偺庬椶 | 棏妱偺條幃 | 棏妱偺夁掱 | 摦暔偺椺 | |||
| 摍 墿 棏 |
|
慡 妱 |
摍 妱 |
俉嵶朎婜傑偱偼丄傎傏摨偠戝偒偝偺妱媴偑弌棃傞 | |
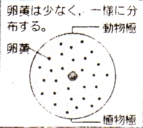
灆旂摦暔 尨嶕摦暔 歁擕椶 |
| 抂 墿 棏 |
|
晄 摍 妱 |
摦暔嬌懁偲怉暔嬌懁偲偱妱媴偺戝偒偝偵堘偄傪惗偢傞 | |
娐宍摦暔 擃懱摦暔 椉惗椶 |
|
|
晹 暘 妱 |
斦 妱 |
摦暔嬌偺泱斦偺晹暘偩偗偱棏妱偑恑傓 | |
嫑椶 唳拵椶 捁椶 |
|
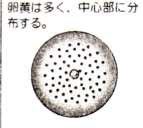
| 怱 墿 棏 |
|
昞 妱 |
暘楐偟偰憹偊偨妀偑棏偺昞柺偵堏摦偟丄偦偙偱棏妱偑恑傓 | |
愡懌摦暔乮峛拵椶丄 峛妅椶乯 |
|
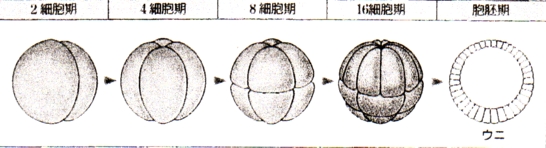
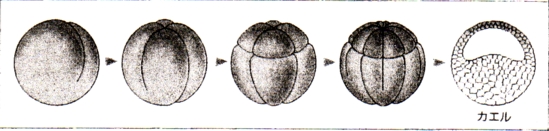
泱偺敪惗
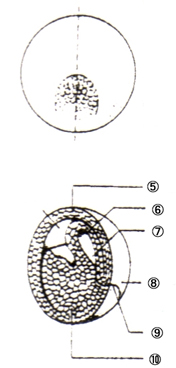
泱丗摦暔偱偼丄棏妱奐巒偐傜敪惗婜乮撈帺偵曔怘傪奐巒偡傞乯傑偱偺屄懱傪巜偡
丒僇僄儖偺敪惗夁掱
 |
庴惛棏丗 丒慡懱偑僛儕乕忬偵曪傑傟偰偄傞 摦暔嬌懁偑崟偄 |
 |
俀嵶朎婜丗 丒摦暔嬌懁偐傜妱傟栚偑擖偭偰丄俀偮偺嵶朎偵側傞 |
 |
俉嵶朎婜丗 丒晄摍妱傪偡傞 丒摦暔嬌懁偺妱媴偺曽偑彫偝偄 |
 |
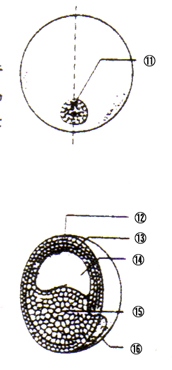
孠幚泱丗 丒棏妱峯偑偁傞 嘆棏妱峯 |
 |
朎泱丗 丒朎泱峯偼摦暔嬌懁偵曅婑偭偰偄傞 嘇朎泱峯 |
 |
尨挵泱弶婜丗 丒泱偺愒摴柺偲怉暔嬌偺拞娫偺晹暘偱娮擖偑婲偙傞 嘊朎泱峯 嘋尨岥 |
 |
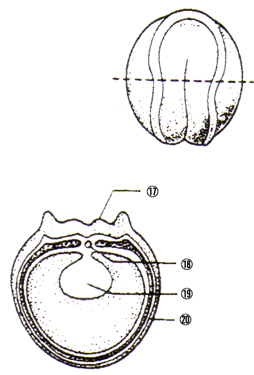
尨挵泱拞婜丗 丒尨岥偼敿墌宍偐傜墌宍偵側傞 嘍奜泱梩 嘐拞泱梩 嘑尨挵 嘒尨岥攚怬晹 嘓尨岥 嘔撪泱梩 |
 |
尨挵泱屻婜丗 嘕棏墿愷 嘖奜泱梩 嘗拞泱梩 嘙尨挵 嘚撪泱梩 嘜棏墿愷 |
 |
恄宱泱慜婜丗 嘝恄宱斅 嘠愐嶕 嘡尨挵 嘢昞旂 |
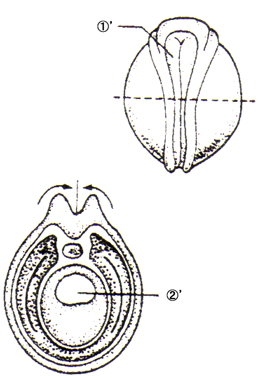 |
恄宱泱拞婜丗 嘆'恄宱峚 嘇'挵娗 |
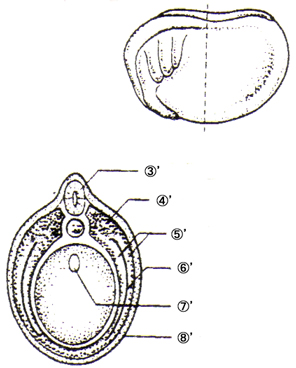 |
恄宱泱屻婜丗 嘊'恄宱娗 嘋'懱愡 嘍'懁斅 嘐'懱峯 嘑'挵娗 嘒'昞旂 |
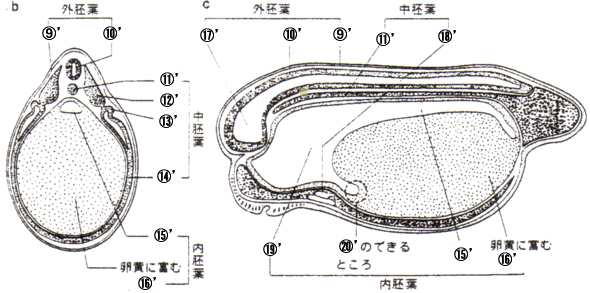 |
旜夎泱丗 嘓'昞旂 嘔'恄宱娗 嘕'愐嶕 嘖'懱愡 嘗'恡愡 嘙'懁斅 嘚'挵娗 嘜'撪泱梩 嘝'擼 嘠'怱憻 嘡'偺偳 嘢'娞憻 |
敪惗偺儊僇僯僘儉
侾俈乣侾俉們倲丗慜惉愢乧乽棏丄惛巕拞偵屄懱偑偁傝丄敪惗帪偵揥奐偡傞乿
侾俉們倲拞婜 丗屻惉愢乧僂僅儖僼偑採彞
丂丂丂丂丂丂丂丂丂丂丂丂丂丂仺僯儚僩儕偺棏傪娤嶡偟偨寢壥丄乽嬒幙側宍懺偐傜條乆側婍姱偑弌尰偡傞乿
挷愡棏偲儌僓僀僋棏
丒挷愡棏乧俀嵶朎婜偵妱媴傪暘棧仺姰慡側泱偲側傞傕偺
丂丂丂丂丂丂 丒偦傟偧傟偺妱媴偑敪惗挷愡傪峴偊傞擻椡偑偁傞
丂丂丂丂丂丂 丒嵶朎暘壔丄寛掕偑斾妑揑抶偄
丂丂丂丂丂丂 丒僂僯丄僇僄儖丄僀儌儕側偳
丂丂丂丂丂丂丂丂丂丂僂僯偺俇係嵶朎婜偵偰丄僂僯偺惓忢敪惗偵偼怉暔敿媴偺彫妱媴偑昁梫晄壜寚
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂仺泱偺敪惗偵偼堦掕偺妱崌偺暔幙乮彫妱媴撪偺暔幙丠乯偑昁梫丠
丒儌僓僀僋棏乧暘棧妱媴偐傜偼晄姰慡側泱偟偐偱偒側偄傕偺
丂丂丂丂丂丂丂丂丂丒嵶朎暘壔偺寛掕偑偼傗偄
丂丂丂丂丂丂丂丂丂丒僋僔僋儔僎乮桳孂摦暔栧乯丄僣僲僈僀側偳
泱偺尨婎暘晍恾乮梊掕堟乯
丂丂丂乧泱偺奺晹暘偑彨棃偳偺傛偆側尨婎傪宍惉偡傞偐傪帵偟偨恾
尨婎乧慻怐婍姱偵暘壔偡傞慜偺嵶朎廤崌懱
僼僅乕僋僩偺幚尡
丒僀儌儕朎泱昞柺晹暘拝怓乮嬊強惗懱愼怓乯
丂丂仺埲屻偺敪惗偱拝怓晹偑壗偵暘壔偟偨偺偐傪挷傋偨
丂丂寢壥丗泱偺梊掕堟乮僀儌儕 尨挵泱弶婜乯

嘆昞旂
嘇懁斅
嘊恄宱斅
嘋愐嶕
嘍愐嶕慜斅
嘐懱愡
嘑尨岥
嘒撪泱梩
宍惉懱偲桿摫
僔儏儁乕儅儞偺幚尡俀乧岎姺堏怉幚尡乮僗僕僀儌儕丄僋僔僀儌儕傪巊梡乯
寢壥丗僀儌儕泱偺奺晹暘偺梊掕塣柦偼丄尨挵泱屻婜偐傜彊乆偵寛掕偟丄恄宱泱弶婜偱偼曄峏偱偒側偔側傞
僔儏儁乕儅儞偺幚尡俁乧擇師泱桿摫幚尡
寢壥丗
丂尨挵泱弶婜偵尨岥攚怬傪棏妱峯撪偵堏怉
丂伀
丂堏怉応強梊掕塣柦偵廬傢偢偵丄愐嶕偵暘壔
丂伀
丂愐嶕傪婎偵恄宱泱丄拞泱梩丄挵娗側偳偑宍惉
丂伀
丂擇師泱偑宍惉偝傟偨
尨岥泱怬偺傛偆偵丄廃埻偺嵶朎偵摥偒偐偗堦掕偺曽岦偵暘壔偝偣傞乮桿摫嶌梡乯傕偺
丂丂丂丂丂丂丂仺宍惉懱乮亖僆乕僈僫僀僓乕乯
桿摫偲楢嵔
丒娽偺宍惉偲楢嵔
丂僯儏乕僐乕僾偺幚尡乮1960偛傠乯乧拞泱梩偵傛傞桿摫
丂丒儊僉僔僐僒儞僔儑僂僂僆傪巊梡
丂丒朎泱拞婜偺梊掕奜泱梩丄梊掕撪泱梩傪暿乆偵庢傝弌偟攟梴
丂丂丂丂丂丂丂丂丂丂丂仺偦傟偧傟偑撪泱梩惈丄奜泱梩惈慻怐偵暘壔
丂丂丂丂丂丂丂丂丂丂丂丂丒偙傟傪枾拝偝偣偰攟梴
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂仺拞泱梩惈慻怐偵暘壔
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂亖梊掕撪嵶朎偑梊掕奜泱梩傪拞泱梩惈慻怐偵桿摫乧拞泱梩惈桿摫
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮偨偩偟丄幚嵺偼孠幚泱傛傝巒傑偭偰偄傞乯
嘆愐嶕
嘇嬝擏
丒摦暔懱乧楢嵔揑桿摫偵傛傝泱奺晹偑暘壔偟丄慻怐丄婍姱偑宍惉偝傟偰偄傞
仭俀侽侽係擭搙侾妛婜婜枛峫嵏
嵶朎暘楐乧嵶朎偼暘楐偵傛偭偰偺傒憹怋
丂丂丂丂丂丂丂丂丂仺堚揱揑側摿挜傪堐帩
暘楐偺庬椶
丒懱嵶朎暘楐丗懱傪峔惉偡傞嵶朎偑憹怋偡傞偨傔偺暘楐
丒尭悢暘楐丗攝嬼巕傪嶌傞偨傔偺暘楐
愼怓懱
丒妀撪偵懚嵼偟丄庡惉暘偼DNA
丒妀撪偵偼晝曽偲曣曽偺愼怓懱偑懚嵼仺憡摨愼怓懱偲偄偆
丒愼怓懱悢偼惗暔庬偵傛傝堦掕
懱嵶朎暘楐偺暘楐夁掱
侾丏娫婜
丒愼怓懱偺庡惉暘偱偁傞DNA偑暋惢偝傟丄攞壔
丒愼怓懱偼僋儘儅僠儞偵側偭偰妀撪偵嶶嵼
俀丏慜婜
丒嶶嵼偟偰偄偨愼怓懱偑嬅廤偡傞
丒拞怱懱偑惎忬懱偵側傞
丒妀枌偑徚幐
俁丏拞婜
丒愒摴柺偵愼怓懱偑暲傇
丒朼悗巺偑摦尨懱偵晅拝偟偰朼悗懱傪宍惉
係丏屻婜
丒愼怓暘懱偑椉懁偵堷偭挘傜傟傞
俆丏廔婜
丒俀屄偺柡妀偑弌棃傞
丒嵶朎幙暘楐偑巒傑傞
怉暔嵶朎偺応崌
丒拞怱懱丄惎忬懱偑側偄
丒嵶朎斅偵傛傝撪懁偐傜奜懁傊偲暘楐偡傞
尭悢暘楐
攝嬼巕偺帩偮愼怓懱悢傪値偲偡傞偲
惛巕丗値亖俀
棏巕丗値亖俀
庴惛棏丗俀値亖係
値亖扨憡
俀値亖暋憡
偡側傢偪尭悢暘楐偼扨憡偺攝嬼巕傪嶌傝弌偡暘楐
尭悢暘楐偺堄媊
丒俀値仺値偵偟側偄偲愼怓懱悢偑憹偊懕偗傞偨傔
丒偄傠偄傠側攝嬼巕偺慻傒崌傢偣偑偱偒丄懡條惈偑偆傑傟傞
尭悢暘楐偺暘楐夁掱
慜婜偱係杮偺攝嬼巕偐傜側傞擇壙攝嬼巕傪宍惉
戞俀暘楐傑偱偁傞
惗怋偲敪惗
惗怋乧惗暔屄懱偑帺屓偲摨偠乽庬乿偺怴偟偄屄懱傪宍惉偡傞偙偲
柍惈惗怋丗懱偺堦晹偑傢偐傟偰怴屄懱傪惗偢傞
丂丂丂丂丂丂丂俙丏暘楐丗侾屄懱偑俀屄懱埲忋偵暘楐偟丄屄懱悢偑憹壛
丂丂丂丂丂丂丂丂丂丂丂丂丂丒扨嵶朎摦暔乧働僀僜僂丄僝僂儕儉僔
丂丂丂丂丂丂丂丂丂丂丂丂丂丒懡嵶朎摦暔乧僀僜僊儞僠儍僋丄僑僇僀
丂丂丂丂丂丂丂俛丏弌夎丗懱偺堦晹偑惉挿丄暘棧偟偰怴屄懱傪惗偢傞
丂丂丂丂丂丂丂丂丂丂丂丂丂丒扨嵶朎惗暔乧僐僂儃乮嬠乯
丂丂丂丂丂丂丂丂丂丂丂丂丂丒懡嵶朎摦暔乧僸僪儔
丂丂丂丂丂丂丂俠丏朎巕惗怋丗恊偺懱偺堦晹偵朎巕宍惉丄敪夎偟怴屄懱傪惗偢傞
丂丂丂丂丂丂丂丂丂丂丂丂丂丒僐僂僕丄僇價丄僉僲僐丄僐働丄僔僟
丂丂丂丂丂丂丂俢丏塰梴惗怋丗怉暔偺塰梴婍姱乮崻丄宻丄梩乯偐傜怴屄懱傪惗偢傞
丂丂丂丂丂丂丂丂丂丂丂丂丂丒僒僣儅僀儌乮夠崻乯丄僕儍僈僀儌乮夠宻乯丄僆儔儞僟僀僠僑乮傎傆偔宻乯丄儀僑僯傾乮梩乯丄儎儅僀儌乮傓偐偛乯
桳惈惗怋丗攝嬼巕偐傜怴屄懱傪惗偢傞
丂丂丂丂丂丂丂攝嬼巕丗崌懱丄愙崌偵娭梌偡傞惗怋嵶朎乮惛巕丄棏乯
丂丂丂丂丂丂丂俙丏椉惈惗怋丗帗梇攝嬼巕愙崌偱愙崌巕乮庴惛棏乯傪偮偔傝丄偦傟偑怴屄懱偵丅
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂愙崌丗摨宆攝嬼巕偺愙崌乧僋儔儈僪儌僫僗丄傾僆儈僪儘
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂堎宆攝嬼巕偺愙崌乧儈儖丄傾僆僒
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂庴惛丗堎宆攝嬼巕偺惛巕偲棏嵶朎偺応崌乧崅摍摦怉暔
丂丂丂丂丂丂丂俛丏扨堊惗怋丗棏偺扨撈敪惗偱怴屄懱乧儈僣僶僠側偳
斏怋曽朄偲堚揱揑摿挜
| 柍惈惗怋 | 桳惈惗怋 | |
| 斏怋 | 憹怋擻棪崅偄 | 憹怋擻棪掅偄 |
| 堚揱揑宍幙 | 恊偲摨偠亖僋儘乕儞 | 椉恊偺慻傒崌傢偣偱堎側傞 |
| 娐嫬曄壔偵懳偡傞揔墳 | 堦斒偵揔墳偼掅偄 乮堚揱揑懡條惈惗偠偯傜偄乯 |
堦斒偵揔墳偡傞壜擻惈崅偄 乮堚揱揑懡條惈惗偠傗偡偄乯 |
尭悢暘楐
桳惈惗怋偲攝嬼巕宍惉丄庴惛
摦暔偺攝嬼巕宍惉偲庴惛
丒攝嬼巕宍惉乧惛巕偺宍惉丗丂丒僑儖僕懱偺撪晹偵愭懱偑偱偒丄妀偺昞柺傪偍偍偆丅
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂拞怱懱偺拞怱棻偺侾偮偐傜慇堐偑偱偒丄旜晹偺拞幉偲側傞丅
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丒妀幙偑嬅弅偝傟偰摢晹傪宍惉偡傞丅拞曅偵偼儈僩僐儞僪儕傾偑娷傑傟傞丅
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂惛巕偑宍惉偝傟傞夁掱偱嵶朎幙偺戝晹暘偼寚棊偡傞丅
丒庴惛乧懱奜庴惛丗懱奜偱庴惛仺惛巕桼棃
丂丂丂丂.丂懱撪庴惛丗懱撪偱庴惛仺棏巕桼棃
丂丂丂丂.丂庴惛棏丗拞怱懱仺惛巕桼棃
丂丂丂丂丂丂丂丂丂丂丂儈僩僐儞僪儕傾仺棏巕桼棃
怉暔偺攝嬼巕宍惉偲庴惛丄敪惗
丒旐巕怉暔偺攝嬼巕宍惉偲庴惛丗偍偟傋懁丂丒壴暡曣嵶朎乮俀n乯
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丒壴暡巐暘巕偺宍惉乮n亊係乯仺暘楐
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丒壴暡娗嵶朎偲梇尨嵶朎偑惗偠傞
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丒梇尨嵶朎偼丄偟偩偄偵壴暡娗嵶朎偺嵶朎暻偐傜棧傟偰偄偔
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂仺壴暡偺姰惉
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 傔偟傋懁丂丒泱偺偆曣嵶朎乮俀n乯
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂仺泱偺偆嵶朎乮値乯
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丒俁夞偺妀暘楐傪峴偭偰俉屄偺妀傪帩偮嵶朎偲側傞
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丒俉屄偺妀偑堏摦偟丄泱偺偆偲側傞
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 俀偮偺庴惛偑摨帪偵婲偙傞仺廳暋庴惛
丒旐巕怉暔偺泱偺敪丗泱乧傗偑偰怴屄懱偵側傞晹暘